
Through the study of malaria, this unit teaches the basics of evolutionary biology and population genetics.
Section 1: Sickle Cell Disease: here students learn about the circulatory system, anemia, alleles, and gene expression.
Section 2: The life cycle of malaria: here students learn about ecological relationships (especially parasitism), prokaryotic vs. eukaryotic cells, disease types, transmission, and vectors.
Section 3: History of malaria: Here students learn how malaria and other diseases evolve. Topics include balancing and directional selection, the trade off between virulence and transmissibility, and disease ecology (specifically, how agriculture changed malaria’s ecology and drove its evolution).
Section 4: Human evolution in response to malaria: Here students learn how to tell when traits are not under selection (using the Hardy-Weinberg equilibrium), how adaptations arise (mutation) and how a selection pressure like malaria can cause evolution (frequency change in alleles). The Sickle cell allele is explained as an adaptation to malaria.
Section 5: Disease ecology. Students will learn why malaria was mostly eradicated in the Northern hemisphere, but not in Africa, why the evolution of resistance makes DDT and other biochemical solutions impermanent, and how public health citizenship should be informed by an awareness of how diseases evolve and spread.
My students do not know much about how malaria or any other disease connects to evolutionary biology. How do pathogens and parasites form? How do humans adapt to selection pressures from disease? How do diseases change in response to our attempts to contain and eradicate them? What are the best ways that society can respond to the challenge of infectious disease? What is the genetic basis of evolution, and how does genetics shape the story of mankind’s war against disease? These are not questions that my average 8th grader is prepared to answer. The problem is not simply that my kids lack knowledge, but also that they think they know things that are not true. I have noticed several persistent myths: Myth 1: When you see variety in a population, natural selection is always at work, making advantageous alleles more frequent. Reality: In fact, most variety in a population involves allele combinations that have no comparative advantage. The alleles that determine hair and eye color, for example, make no difference to fitness. These alleles maintain a stable frequency precisely because they are not under selective pressure. Underlying issue: Many kids are not able to distinguish between traits that are under selective pressure and ones that are not. Most do not know that there is such a distinction to be made. Nor do they grasp that selection pressures can take different forms in different environments. The sickle cell allele, for example, is under negative selection here in the United States (where it gives no protective benefits) but is stable in sub-Saharan Africa (where it offers some protection against malaria). Myth 2: Evolution happens over a time scale of millions of years, so nothing we do is going to cause other species to evolve within our lifetimes, nor is anything that happens to us going to cause us to evolve. Evolution just does not work that fast. Reality: If a trait is under selection, evolution can be rapid. In the 20th century, mosquitoes evolved resistance to several kinds of pesticides. Malaria has fully adapted to what was once mankind’s frontline anti-malarial, chloroquine, and is fast overcoming our current drug, artemisinin. Human beings evolve more slowly, but they too have changed. Parasites and pathogens exert selection pressures on human populations, and these pressures have resulted in human evolution (albeit at a pace far slower than that of malaria or mosquitoes). Duffy negativity and Sickle Cell are two examples of human adaptation due to selection pressure from malaria. Underlying issue: Many kids do not understand how rapidly pathogens, parasites and vectors develop resistance, so they don’t understand why antibiotics are losing effectiveness, or that insecticides are just temporary solutions to problems of vector control. Myth 3: There are two alleles for every trait, one that is dominant and expressed in phenotype, the other recessive and not expressed. Reality: In fact, codominance is far more typical. Heterozygote carriers of the sickle cell trait, for example, have a selective advantage over homozygotes (in malarial areas) because both of their hemoglobin alleles express. Underlying issue: This myth is a natural generalization of the simple Mendelian genetics my kids have been taught. Since Mendel’s peas had traits that were clearly dominant or recessive, my kids conclude that all alleles work that way. This fogginess about gene expression leads to misunderstandings about selection. They wrongly conclude that only the dominant allele gets expressed in phenotype, so only the dominant allele comes under selection pressure. This misunderstanding impedes their ability to understand how humans and other organisms evolve. These mistaken assumptions are problematic in part because they preclude informed public health citizenship. Good health citizenship involves many dimensions, such as activism for clean water, clean air and food safety. But it also involves a consciousness of how diseases form and change and how to respond to disease in an intelligent way. First, students need to know that when we use pesticides or medicines, we are exposing the organisms responsible for disease to powerful selective pressures, and these pressures will cause rapid directional selection. Second, they need to grasp that a disease reservoir anywhere is a threat to people everywhere, particularly since global warming will likely expand the reach of many diseases, including malaria. Air travel and globalized commerce ensure that an emergent disease will have a good chance to spread to other nations before it can be recognized and contained. Because disease management is a global challenge, kids need to understand that support for global institutions like the WHO is not just a hand-out to impoverished nations, but enlightened self-interest. Lastly, kids need to grow into adults who support systemic solutions to the problems of disease, not just the temporary fixes provided by pesticides and medicines. The history of malaria shows that nations that beat malaria did so through social development, by improving their schools, primary health facilities, housing, and infrastructure (especially drainage). Malaria and mosquitoes are very adaptable, but there are challenges that even they cannot overcome. Malaria cannot spread without mosquitoes, and mosquitoes cannot breed without standing water or feed on people who sleep in well-built houses. The failure of malaria eradication in Africa and the success of such efforts in wealthier nations teach a simple lesson, that without basic economic reforms and the political and financial commitments that make such reform possible, there is no lasting solution to the problem of disease. Malaria is a useful topic for 8th graders to study. It can be used to teach a wide range of science standards. It connects science and social studies since the ecology of malaria requires a political and historical story along with a scientific one. It can be used to teach some of the key middle years math standards. The study of selection pressures, frequency changes among alleles, and disease mortality and morbidity provide real-world application for the basic concepts of probability and statistics. Lastly, malaria has a human face that makes it intrinsically interesting to a young teenager. Videos, interviews and pictures make the burden of this disease vivid and real, and hopefully this will stimulate self-guided learning. A study of malaria can also help to create a basis for informed public health citizenship. Public health citizenship requires a suspicion towards easy biochemical fixes, a sense of global responsibility, and support for the broad economic reforms that provide permanent solutions to disease. These dimensions of citizenship are obvious once one understands the evolutionary biology of disease, and a study of malaria can teach kids this biology in a way that is immediate and real. The word ‘malaria’ comes from the Latin words for ‘bad air.’ For centuries people noticed that proximity to swamps accompanied higher rates of infection, so they believed that foul smells from swamps (miasmas) were responsible for the disease. What they did not know is that malaria is caused by a parasitic protozoan called a plasmodium. Alphonse Laveran, a French physician, discovered this in 1880, when he studied the blood smears of patients who had died of malaria in Algeria.[1] Squirming within their red blood cells were thousands of single celled protozoans, feeding and reproducing. A plasmodium is very different from a bacterium or a virus. Since it is a eukaryote, a plasmodium has a nucleus to contain its large and complicated genome. Where bacteria have only a single chromosome, the malaria plasmodium has fourteen. On these fourteen chromosomes are 23 million base pairs, encoding approximately 5300 genes. In contrast to ordinary, ‘free living’ (non-parasitic) protists, a large proportion of plasmodium’s genes are dedicated to helping it to grow and reproduce within the bodies of its two hosts.[2] Those two hosts are the female Anopheles mosquito, which malaria does not seem to hurt, and vertebrates like humans, who suffer serious harm and often death as a result of infection. Thanks to its complex genome, the malaria plasmodium can perform an incredible series of transformations, each of which is necessary for the continuation of its life cycle. Malaria enters the bloodstream of a human host through a mosquito bite. When the mosquito bites, she injects saliva to prevent the blood from clotting, and within this saliva is the malaria plasmodium. Malaria rides the blood stream to the liver, where it enters a Kupffer cell. The job of a Kupffer cell is to filter harmful substances out of the bloodstream, like cellular debris and dead blood cells. But malaria, thanks to the sharp, needle-like form it has at this stage (the ‘sporozoite’ phase), punches through the Kupffer cell and enters the tissue of the liver.[3] There, within the infected liver cell, it becomes a ‘schizont’, performing thousands of asexual nuclear divisions. Eventually the infected liver cell explodes, and thousands of copies of the parasite (in this form it is called a ‘merozoite’) flood the bloodstream. The merozoite has special structures that allow it to penetrate red blood cells. Within the blood cell, the parasite feeds and reproduces. Soon, the blood cell ruptures, putting yet more parasites into the bloodstream. If a mosquito bites a person who is infected with malaria, the parasite will enter the mosquito along with her blood meal. The cooling of the blood in the mosquito’s stomach triggers yet another transformation of the plasmodium. Dormant cells called gametocytes change into eggs and sperm and reproduce sexually. Fertilized eggs make their way to the stomach lining of the mosquito, where they transform into cysts. Each cyst makes thousands of sporozoites, which travel to the mosquito’s salivary glands. There, the parasite is ready to ride the mosquito’s saliva into its next vertebrate host. Malaria is an ancient disease. The plasmodium has been found in the belly of a midge preserved in amber 100 million years ago, suggesting that malaria fed on birds and reptiles long before the evolution of man.[4] For as long as human beings have kept written records, they have documented the effects of malaria. Ancient Egyptians recorded fevers and other symptoms that we can recognize as effects of malaria. They also knew that malaria increased in proximity to swamps and marshes.[5] Before the late 20th century, European doctors recognized that the disease appeared in different forms. There was the “benign tertian” form, with relatively mild fevers appearing every three days, and the “malignant quartan” form, which was far more deadly and returned every fourth day.[6] They did not know it, but what these doctors were seeing were the different reproductive cycles of two distinct species of plasmodium. The ‘tertian’ form was the older, less virulent species of the plasmodium, P. vivax, while the ‘quartan’ was the newer and far deadlier P. falciparum. The one constant in the history of malaria has been its astonishing power to kill. If you counted its total victims over the centuries, that number would be in the billions. Most of the dead today and throughout history are African children under the age of 5. But malaria has not always been a tropical disease. It used to be endemic to the southern states of the U.S. and to southern Europe, particularly Italy and Greece. In 1900, there were about 80,000 deaths per year in Europe and North America from malaria, but by 2000, there were less than 20. Africa’s story has no similar happy ending. After peaking in the 30’s at 3.5 million deaths per year, malaria deaths in Africa declined to about a half million per year by the 70’s. Then, they steadily rose again, reaching more than 1 million per year by the year 2000. Then there was a second decline between the year 2000 and 2016, with Africa’s death rate falling to 413,000 in 2016.[7] In the last few years, however, progress against the disease has stalled or reversed.[8] These fluctuations in mortality raise a host of questions. Why was malaria eliminated in Europe and the United States, but not in Africa? Why did mortality in Africa fall until the 70’s and then rise again? Why has progress in Africa stalled since 2000? The causal explanations to these questions are complex and cannot be reduced to a single factor, but according to Randall Packard, there is one particularly important distinction. In Europe and North America, the elimination strategy was ecological, while in Africa, the strategy has been mainly biochemical. Europe and North America changed the way that people lived, reducing malaria’s habitat. In Africa, the strategy has been to attack the vectors of the disease, and while this approach yielded great short-term gains, it was eventually undermined by the evolution of resistance among both mosquitoes and the malaria plasmodium.[9] In the United States, malaria was in decline long before the advent of DDT after WWII. Most of these declines were due to capital improvements in agriculture and infrastructure and to vast improvements in the living conditions of agricultural workers. Modern sanitation and drainage systems came to the large cities of the Northeast (Boston, New York, Philadelphia) in the first two decades of the 20th century, greatly reducing malaria in those areas. Accelerating industrialization in the North, combined with capital improvements in agriculture, especially the replacement of farm laborers with machines, led to a decline in rural population and a consequent decline in malaria mortality. In the South, agricultural feudalism (a small land-owning class with political control over a large, impoverished labor force) persisted long after the end of slavery, and this allowed malaria to endure. Labor was cheap, which removed the incentive for capital investment, and there was little political push to attack malaria since the people who suffered from it were (mostly) African-American laborers, denied suffrage under the South’s Jim Crow laws. Things began to change with the arrival of the Boll weevil. This beetle, which feeds on the buds of cotton plants, led to major declines in cotton cultivation in the South.[10] With a shrinking agricultural labor force, malaria mortality also fell. The arrival of the New Deal in the 30’s dealt another heavy blow to malaria. In the Tennessee Valley, the TVA made major improvements to the living conditions of poor workers. Electricity from new dams allowed industrialization and made farms more productive. Housing was modernized and windows were screened. Vector control was also utilized, but the key factor in success was the change in the way people lived and worked. Due to these changes, malaria had “all but disappeared from the TVA by 1938.”[11] Italy in the 19th century was in some ways like the American South. Here, too, there was a small landowning class with little incentive to make capital improvements or improve living conditions for workers. In 1922, the fascist government of Benito Mussolini came to power, and unlike previous administrations, which had largely served the interest of the land-owning class, Mussolini was determined to uplift Italy’s poor. His ‘bonification’ programs created dams and drainage systems, rural schools and primary care clinics. Better housing, better education and higher incomes changed the way people lived, and this, along with capital and infrastructure improvements, led to a steep decline in mortality from malaria.[12] Success against malaria in Europe and the U.S. came from the combination of vector control and structural economic reform. In post-WWII Africa, however, there was a singular focus on vector control with little attempt to improve basic living conditions. This choice was not made by Africans, but rather by the wealthy European and American nations who controlled policy at the WHO. Those nations were moving to the political right, becoming more conservative as a result of their conflict with the Soviet Union. “With this shift,” writes Packard, “leaders began to view models of social medicine that linked health to broader social and economic reform as being influenced by socialism and thus politically suspect.”[13] Few scientists opposed this policy because most were confident that their new, biochemical arsenal was by itself sufficient to eradicate malaria. DDT had achieved some spectacular successes, including the elimination of malaria from much of Eastern Europe and the Caribbean, and many thought they could repeat this success in Africa. At first, it looked as if they might succeed. Malaria deaths fell dramatically through the 50’s and 60’s. In the 70’s, this trend reversed. DDT was abandoned because of its devastating environmental effects and increasing resistance among mosquitoes. Chloroquine, a cheap front-line anti-malaria therapy, also had to be abandoned because P. falciparum had become resistant to it. Alternative drugs and pesticides were far more expensive. By 1969, it was evident that vector control had reached the limit of its effectiveness, and the WHO officially abandoned its Eliminate Malaria Program. Funding for anti-malaria programs was greatly diminished.[14] At the same time, Africa’s population was growing exponentially while its GDP steadily fell. Per capita GDP in Africa decreased by about 45% between 1960 and 1999, while mortality due to malaria grew by 60% during this same time period.[15] This was no coincidence. Poverty and malaria are mutually reinforcing; malaria causes poverty, but poverty also causes malaria. African nations hoped that by large-scale borrowing they could reduce poverty and malaria, but their efforts to industrialize were mostly unsuccessful. This added crushing levels of debt to their woes. In the early 80’s, HIV arrived. This new epidemic consumed scarce medical resources that might otherwise have gone into fighting malaria. Researchers also learned that an immune system compromised by HIV was far less able to combat the effects of malaria.[16] War is also an exacerbating condition of malaria. As Africa’s poverty and population increased, its wars became more frequent and severe.[17] All these factors—war, HIV, debt, international disinterest, and resistance to drugs and insecticides—combined to greatly increase malaria mortality between 1970 and 2000. The main reason for the dip in malaria mortality after 2000 was the development of a new and powerful anti-malarial drug, artemisinin. But the decline in malaria mortality has since levelled off. Artemisinin has grown less effective. Like chloroquine, which was effective until the plasmodium developed resistance in the 1950’s and 60’s, today’s artemisinin-based therapies are not working as well as they used to. Failure rates for these drugs are particularly high in the Mekong delta (Cambodia, Thailand, Laos)[18] and while these mutations have not yet spread to Africa, they are expected to eventually.[19] Another reason for the stall in progress is pesticide resistance among mosquitoes. Malathion was an expensive but effective replacement for DDT, but mosquitoes have become increasingly resistant to malathion and most other available insecticides.[20] Malaria’s disease reservoir (species in which it can continue its lifecycle) is enormous, with 200 species of plasmodium affecting every major group of land vertebrates. Malaria that affects one species can jump to another, given the right mutations. And malaria is highly mutable, with multiple polymorphisms on hundreds of different genes. This makes malaria extremely resilient against eradication efforts.[21] Although some still believe that we will use our technology to eliminate malaria, as smallpox was once eliminated, Pedro Alonso, director of WHO’s malaria response, is less optimistic. “With the tools we have today,” he says, “it is most unlikely eradication will be achieved.”[22] One piece of good news is that there is a high percentage of acquired immunity in areas where malaria is endemic, which means that if the disease does not kill you as a child, it is unlikely to when you are an adult. This is also why children under 5 represent 67% of malaria’s victims.[23] Their immune systems are overwhelmed before they can learn to combat the disease. Ironically, this acquired immunity has become a political obstacle to more vigorous attempts at eradication, and even to the widespread use of a simple implement like an insecticide-treated bed net. People who are used to living with malaria treat it as ordinary and unavoidable and are therefore reluctant to devote scant resources to its elimination, or even to go through the bother of putting up a net each night.[24] This “malaria culture,” together with increasing resistance in the mosquito and the plasmodium, make it likely that malaria will continue to be with us for a very long time. How does malaria kill? Malaria hides from the body’s immune system in the cells of the liver, where it quietly reproduces. Periodically (about every three days, depending on the species of plasmodium) these cells rupture, flooding the blood stream with parasites. It is during this stage that an infected person suffers fever, as the immune system responds to the sudden influx of contaminants from the ruptured blood cells. Victims suffer severe aches, respiratory distress, and fatigue as the destruction of red blood cells deprives their bodies of oxygen. Fever and anemia come with every strain of malaria, but if one is infected with Africa’s most common strain, P. falciparum, there is an additional threat. Infected red blood cells are recognizable by the body’s spleen, where they are filtered out and destroyed, but P. falciparum has evolved a defense against the spleen. It causes changes to the surface area of the infected red blood cell, making it stickier and therefore more likely to adhere to the sides of veins and arteries. A blood cell that gets stuck will not be processed by the spleen. These sticky blood cells clog circulation in the major organs, including the brain, leading to strokes and organ failure.[25] Because malaria has killed so many people for so long, it has caused adaptive changes in the human genome. A mutation that gave resistance to the plasmodium would spread rapidly in populations where morbidity (the frequency of infection) was high. In the 1940’s, the geneticist J.B.S. Haldane observed that disorders of the blood like Sickle Cell disease and Thalassemia were common in regions where malaria was endemic. He formed the hypothesis that malaria was the selection pressure responsible for these disorders.[26] Haldane’s student, Anthony Allison, would later confirm his teacher’s hypothesis. Anthony tested thousands of Africans for the presence of the sickle cell allele. He found that “among tribes living close to the coast of Kenya or to Lake Victoria, the frequencies exceeded 20%, whereas among several tribes living in the Kenya highlands or in arid country, the frequencies were less than 1%. These differences cut across linguistic and cultural boundaries and were independent of blood group markers.”[27] This data supported the hypothesis that sickle cell was an adaptation to combat malaria. In the dry highlands, where the Anopheles mosquito cannot live, frequency of the allele was very low. In the wet, warm lands around Lake Victoria, where mosquitoes could carry the plasmodium from host to host, the frequency of sickle cell heterozygotes (possessing one copy of the sickle cell allele and one copy of the normal allele for hemoglobin) was as high as 40%. To confirm that he was looking at causation, not just correlation, Anthony then tested thousands of children under the age of four. He ignored older children because their acquired immunity to malaria made it impossible to tell if they received any benefit from having the sickle cell gene. Among younger children, who had no acquired immunity, those who were heterozygous for the sickle cell allele (AS) showed far lower parasite counts than those with two copies of the dominant allele (AA). Given the established link between parasite counts and mortality, Anthony concluded that “AS children are more likely to survive through early childhood in a highly malarial environment than AA children.” But how did the sickle cell allele confer resistance to malaria? In any red blood cell, filaments of actin work as a cytoskeleton. P. falciparum hijacks these actin filaments and uses them to move its own proteins to the cell surface. One of these proteins is adhesin, which makes the surface of the blood cell sticky. This stickiness causes the red blood cell to stick to the vascular walls and this is beneficial for the parasite because it protects infected cells from filtration by the spleen. In cells with the sickle cell allele, however, actin is not able to polymerize into long chains. This prevents malaria from moving adhesin to the cell’s surface, and without this sticky protein, infected cells cannot stick to vascular walls and are thus more easily destroyed by the spleen.[28] Unfortunately, this resistance comes at a price. People who are homozygous for the sickle cell allele start to manifest symptoms of Sickle Cell Disease (SCD) when they are about five months old. Their sickle shaped red blood cells are rigid and this rigidity prevents them from flowing into small capillaries. The result is oxygen starvation in the body’s tissues. This causes severe pain, damage to the major organs, and strokes. Also, sickled cells have a much shorter life span than normal red blood cells. They live for 10-20 days, versus 90-120 days for normal cells. The body cannot replace these short-lived cells quickly enough, resulting in severe anemia, further reducing the circulatory system’s ability to carry oxygen. There is no cure for SCD, other than a bone marrow transplant, and these transplants are often unsuccessful.[29] Heterozygotes (AS) are said to have the sickle cell trait, but not SCD. Usually they do not suffer from anemia, but, as Anthony Allison discovered, they do enjoy some immunity to malaria as compared to homozygotes (AA). In areas where malaria is endemic, heterozygotes have a higher fitness than either homozygote because AA homozygotes have no protection from malaria while SS homozygotes suffer from Sickle Cell Disease. In areas free from malaria, however, the AA homozygote has the highest fitness. About 100,000 Americans suffer from SCD (.033%). Most are African Americans descended from African populations where malaria gave a selective advantage to the AS genotype. Since here in the United States there is no fitness advantage to the AS genotype (and an enormous fitness cost to the SS genotype), it is presumed that the frequency of the sickle cell allele will eventually decline.[30] Carriers of the gene (those with sickle cell trait) are largely asymptomatic, however, and since this allows the sickle cell allele to ‘hide’ in heterozygotes, one can expect this decline to be very slow. The emergence of the sickle cell allele was not humanity’s first adaptation to malaria. An earlier mutation, the Duffy negative blood group, appeared tens of thousands of years ago and protected Africans from what was then the dominant malarial strain, P. vivax. By the 1930’s it was well-known that the majority of African Americans were immune to P. vivax. In the 50’s, researchers discovered that most were also Duffy negative, which is “extremely rare in other racial groups without black admixture.”[31] To establish a causal link between the immunity and the blood group, researchers allowed P. vivax infected mosquitoes to feed on groups of prisoners drawn from two prisons in the United States, the Maryland House of Correction in Jessup and the Georgia State Penitentiary in Atlanta. “Of the 11 blacks and six whites challenged in the six studies, erythrocytic infection developed in six blacks and all six whites. Five blacks never had a positive blood film for P. vivax during the subsequent three to six months of daily examinations. The only consistent difference between erythrocytes of blacks resistant and susceptible to P. vivax was in the Duffy blood group system.” These results showed that the Duffy blood group was indeed an adaptation that gave many African-Americans immunity to P. vivax.[32] A person who is Duffy-negative does not produce a protein that is typically found on the surface of the red blood cell. That protein, called the Duffy-Antigen Chemokine Receptor (DARC) binds to chemicals that are secreted by cells when they are inflamed. This increases blood flow to inflamed tissues. But DARC is also the binding site for P. vivax. Without DARC, P. vivax is unable to enter the red blood cell and its life cycle is interrupted.[33] But why would Africans and their African-American descendants have immunity to a strain of malaria that is not endemic to Africa? The two main species of malaria, P. falciparum and P. vivax, have different geographical distributions. P. falciparum is the most common strain in Africa, while P. vivax is the dominant variety in Central and South America and Southeast Asia. Once it was thought that P. vivax had emerged and evolved outside of Africa, but studies of mitochondrial DNA in apes suggest that in fact, P. vivax is of African origin, and traveled with early humans as they migrated out of Africa. A strain of P. vivax is still endemic to populations of African apes. Researchers discovered that the genome of this strain was 98% identical to the P. vivax that infects humans and inferred that the two strains have a relatively recent common ancestor. But human strains of P. vivax are far less diverse than those found in African apes. This suggests that at some point in history there was a genetic ‘bottle neck,’ a movement of a small sample of the population of P. vivax out of Africa. Because all the current strains of P. vivax descend from this small sample, they have less genetic diversity than the variety found in African apes.[34] This explains why the Duffy negative blood group is so common in Africa, where P. vivax is currently rare. “The P. vivax ancestor likely infected both humans and apes in Africa before being eliminated in humans there by the spread of the Duffy-negative mutation. The current human-infecting parasites (P. vivax in Asia and the Americas) represent a lineage that escaped out of Africa.” The Duffy negative group is of little use to Africans today, as it has no effect on P. falciparum. It may be vestigial, like wisdom teeth or the appendix, or it might someday protect Africans from the primate vivax strain, if that strain developed a mutation that allowed it jump to humans. About ten thousand years ago, P. falciparum evolved to exploit the niche left vacant by the disappearance of P. vivax. P. falciparum has a relatively low level of genetic diversity, compared to other malarial strains, and this suggests another ‘bottle neck’ event, not unlike the movement of P. vivax out of Africa. In this case, the parasite moved from gorillas to humans, causing a founder effect. Genetic diversity among P. falciparum remains low because colonization happened recently, and because the number of colonizing ancestors was small.[35] It is no coincidence that the P. falciparum emerged at the same time as agriculture, nor that it was so much more virulent than the earlier P. vivax. Agriculture creates perfect mosquito breeding grounds and puts large populations of humans in close contact to one another. P. vivax has a low level of virulence because, at the time of its emergence, human populations were low and widely scattered, which meant that the parasite had to insure the survival of its host for long periods in order to come into contact with a new host. P. falciparum lives in agricultural communities with vast numbers of hosts readily at hand, thus has no similar selection pressure towards lower virulence. In a pathogen population, competition between individuals creates a selection pressure towards greater virulence, if one assumes that the greater the parasite load, the greater the chance of transmission. This pressure is balanced by the need to keep the host alive long enough to spread to the next host, which tends to reduce virulence. In P. vivax, that balance point favors low virulence. This, along with vivax’s ability to survive winters by remaining dormant within the cells of the liver, suggests that it evolved to parasitize humans that would only sporadically encounter one another. P. falciparum, by contrast, has evolved a much higher virulence, probably because the large, sedentary communities created by agriculture made it possible for the parasite to move from host to host much more quickly.[36] This, in turn, explains why the sickle cell allele is so common in areas where P. falciparum is endemic. P. falciparum’s extreme virulence creates a powerful selection pressure in favor of the sickle cell allele. The long history of humanity’s war with the malaria plasmodium is an example of the Red Queen hypothesis. That hypothesis states that an organism must keep evolving to maintain its niche, because its competitors (or predators, or parasites, or hosts) are also evolving. Our current use of medical and chemical technology to combat malaria is in some ways like our earlier evolution of the Duffy negative blood group. Each provided a temporary respite from the parasite, but by constricting the parasite’s niche, each also created selective pressures that caused it to change into new, more resistant and more virulent forms. P. falciparum has not yet evolved a way to defeat the sickle cell allele, but since this allele reduces parasite load, it also creates a selective pressure. This selection pressure may someday drive further adaptation in malaria, although this remains to be seen. What is certain is that malaria will continue to evolve in ways that improve its ability to parasitize its vertebrate hosts, just as those hosts, including humans, will evolve new ways to resist this parasitism. That does not mean we must wait passively for a benign mutation to free us from this disease burden, as Duffy negativity once did. Malaria can be defeated by other means and its elimination in the developed world proves this. Whether we will embrace the expensive ecological strategies that enabled these victories or remain wedded to cheaper, impermanent biochemical fixes, is another open question. [1] Nobel Prize Biography of Alphonse Laveran. https://www.nobelprize.org/prizes/medicine/1907/laveran/biographical/ [2] “Compared to the genomes of free-living eukaryotic microbes, the genome of this intracellular parasite encodes fewer enzymes and transporters, but a large proportion of genes are devoted to immune evasion and host–parasite interactions.” Gardner, Malcolm et al. “Genome Sequence of the Human Malaria Parasite Plasmodium Falciparum.” Nature, Oct. 2002. [3] Barnwell, John. “Hepatic Kupffer Cells: The Portal That Permits Infection of Hepatocytes by Malarial Sporozoites?” Hepatology, May 2001. [4] Podcast: “This podcast will kill you: Malaria”. http://thispodcastwillkillyou.com/2018/02/10/episode-8-abracadabra-go-away-malaria/ [5] Fagan, Toby. “When was malaria first discovered and by whom?” Scientific American, July 17, 2000. [6]Carter, Richard. “Evolutionary and Historical Aspects of the Burden of Malaria.” Clinical Microbiology Reviews, Oct. 2002. [7] World Malaria Report, World Health Organization, 2018, p. 41. [8] “Although there are some bright spots in the data, the overall decline in the global malaria burden has unquestionably levelled off. And, in some countries and regions, we are beginning to see reversals in the gains achieved.” WHO news release, Nov. 2017. https://www.dw.com/en/world-health-organization-malaria-treatment-stalls-as-funding-flatlines/a-41577312 [9] Packard, Randall. Malaria: The Making of a Tropical Disease. Johns Hopkins Univ. Press, 2007: “Efforts to control malaria since the end of the 19th century have been largely driven by a narrow vision of the disease and its causes that has privileged biological processes and focused on attacking mosquitoes and malaria parasites. This biological model has informed public health policy since the discoveries of Laveran, Ross and Grassi. Put simply, malaria policy has largely ignored the human ecology of malaria. The failure to link ecology and policy has prevented the elimination of malaria as a serious public health problem in many areas of the globe” p. 247. [10] In Georgia, for example, cotton cultivation went from 5.2 million acres in 1914 to 2.6 million in 1923. Packard, p. 77. [11] Packard, p. 133. [12] Packard, p. 128. [13] Packard, p. 145. [14] Total spending went from 1.4 billion between 1957-1967 to 250 million between 1967 and 1977. Packard, p. 169. [15] Packard, p. 205. [16] https://www.who.int/malaria/areas/high_risk_groups/hiv_aids_patients/en/ [17] The First and Second Congo Wars, also called Africa’s World Wars, took place between 1996 and 2003, and involved many of the nations where malaria infection is endemic: DRC, Angola, Uganda, Rwanda, Burundi, etc. [18] WHO World Malaria Report, 2018: In People’s Democratic Republic of Laos treatment failure rates of 10%, 14.3% and 17.2% were observed in 2013, 2014 and 2017, respectively. [19] WHO status report: “Artemisinin and artemisinin-based combination therapy resistance.” Oct. 2016. [20] “Resistance to the 4 commonly used insecticide classes – pyrethroids, organochlorines, carbamates and organophosphates – is widespread in all major malaria vectors across the WHO regions of Africa, the Americas, South-East Asia, the Eastern Mediterranean and the Western Pacific.” WHO news release, Feb. 2020. https://www.who.int/malaria/areas/vector_control/insecticide_resistance/en/ [21] Kwiatowski, Dominic. “Malaria genomics: tracking a diverse and evolving parasite population.” Lecture on youtube: https://www.youtube.com/watch?v=bb1lgvx3HPg&t=295s [22] “Malaria Eradication Not Currently Possible.” Physician’s Weekly, Aug. 2019. [23] WHO Malaria: Key Facts. https://www.who.int/news-room/fact-sheets/detail/malaria [24] Shah, Sonia. “Three reasons we still have not gotten rid of malaria.” Ted talk, Sept. 2013: https://www.youtube.com/watch?v=SF242mxtZN8 [25] “How Malaria Kills.” Medecins Sans Frontiers website: https://www.msf.org/how-malaria-kills [26] Sabeti, Pardas. “Natural Selection: Uncovering Mechanisms of Evolutionary Adaptation to Infectious Disease.” Nature Education, Jan. 2008. [27] Allison, Anthony C. “The discovery of resistance to malaria of sickle‐cell heterozygotes.” Biochemistry and Molecular Biology Education, Nov. 2006. [28] Cyrklaff, Marek et al. “Hemoglobins S and C Interfere with Actin Remodeling in Plasmodium falciparum–Infected Erythrocytes.” Science, Dec. 2011. [29] National Institute of Health (NIH) website: https://ghr.nlm.nih.gov/condition/sickle-cell-disease#genes [30] Futuyma, Douglas. Evolutionary Biology, 3rd Edition, 1998: p. 385. [31] Miller, Louis, et al. “The Resistance Factor to Plasmodium vivax in Blacks — The Duffy-Blood-Group Genotype, FyFy.” New England Journal of Medicine, Aug. 1976. [32] In malaria-endemic areas of Africa, the frequency of Duffy negativity is extremely high (between 95%-100%). But Duffy negativity is less frequent in the United States, mainly due to interracial sex between masters and slaves: “One of the many human costs that were borne by African-American communities during slavery was that interracial sex between master and slave resulted in a loss of Duffy negativity, and a larger percentage of the U.S. black population became susceptible to vivax infection.” Webb, James. Humanity’s Burden: A Global History of Malaria. Cambridge U. Press, 2009, p. 90. [33] National Center for Biotechnology Information website: https://www.ncbi.nlm.nih.gov/books/NBK2271/ [34] Loy, Dorothy et al. “Evolutionary history of human Plasmodium vivax revealed by genome-wide analyses of related ape parasites.” Proceedings of the National Academy of Sciences, Aug. 2018. [35] Loy, Dorothy, et al. “Out of Africa: origins and evolution of the human malaria parasites Plasmodium falciparum and Plasmodium vivax.” International Journal for Parasitology, Feb. 2017. [36] “The change in lifestyle from small nomadic groups to larger settled communities and the subsequent increase in population size would have, for the first time, provided conditions capable of sustaining P. falciparum transmission… P. falciparum, with its high pathogenicity and short term survival in the host, would have faced a high risk of extinction in small, dispersed groups of hunters and gatherers.” Hume, Jennifer et al. “Human migration, mosquitoes and the evolution of Plasmodium falciparum.” Trends in Parasitology, March 2003.Problem Statement
Rationale
Background: Content Knowledge
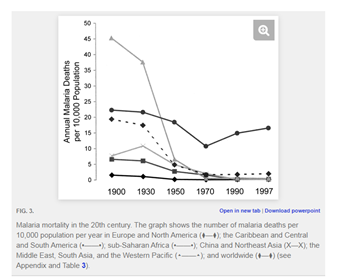
My unit plan begins by introducing students to victims of Sickle Cell Disease. This will hopefully raise questions that the rest of the unit will answer: what causes SCD, and why are most Americans who have it of African descent? The rest of the unit aims to answer those questions. We begin by studying the life cycle of malaria, then look at the evolution of malaria in response to human adaptation and technological change. Then, the heart of the unit: an introduction to population genetics and how selection pressures like malaria drive human evolution. Lastly, we examine mistakes in our efforts to eradicate malaria, with a focus on the evolution of resistance. The aim here is to raise a question fundamental to public health citizenship: how do we respond to this disease (or any disease) in a way that is informed by an understanding of evolution?
Objective: The aim of this introductory lesson is to connect students to people who suffer from SCD, and help students understand what causes their symptoms. This lesson should also be used to teach or review basic facts about the circulatory and respiratory system. Use lecture (direct instruction) to connect the symptoms of SCD to the defective hemoglobin gene, emphasizing several key concepts: gene expression, cellular respiration, the circulatory and respiratory systems. Kids should understand what anemia is, how it affects a patient’s life, and how it results from SCD. Ask, but do not answer, the question of why almost all patients with SCD are of African descent. Hopefully this excites their curiosity and will deepen their interest in subsequent lessons in this unit. Key vocabulary: diagnosis, symptom, anemia, red blood cells, bone marrow, hemoglobin, allele, gene expression, protein, cellular respiration, stroke, bone marrow, transfusion Sequence: First, view a couple of videos where victims tell their stories: Video 1: Alexandria’s story: https://www.youtube.com/watch?v=2CsgXHdWqVs This video includes a simple explanation of sickle cell anemia, with a focus on a young lady who suffers from the disease. Video 2: Teonna’s story: https://www.youtube.com/watch?v=xWLjeaWyGps After videos, students work in small groups. Using the Mayo clinic website and their notes on the video, the groups define vocabulary and answer three big questions on the graphic organizer: causes (focus on gene expression) symptoms (focus on anemia), treatments (focus on bone marrow transplants and transfusions). Mayo clinic overview of disease: https://www.mayoclinic.org/diseases-conditions/sickle-cell-anemia/symptoms-causes/syc-20355876 Standards: S8.B.1.1.1 Describe the structures of living things that help them function effectively in specific ways (e.g., adaptations, characteristics). S8.B.2.1.3 Explain that mutations can alter a gene and are the original source of new variations. Objective: Key vocabulary: infectious disease, parasite, host, symbiosis, eukaryote, mitosis, meiosis, asexual reproduction, sexual reproduction, protist, life cycle, fungi, incubation period, reproductive rate, mortality (case fatality), zoonotic disease, emerging diseases, vector, transmission, tropical disease Sequence: After viewing, kids could work on a four column organizer in small groups: the four columns are bacteria, viruses, parasites, and fungi. On the y axis of this organizer are two rows: First row: What is it? (here students give general descriptions of each category). Second row: Example? (Here students briefly explain an example of a disease caused by each kind of organism.) After viewing, students work in small groups to complete an organizer on the types of ecological relationships: predator-prey, symbiosis. Break down symbiosis into parasitism, mutualism, and commensalism. Have boxes for kids to explain each type of relationship and provide an example of each. Part two, mosquito host: https://www.youtube.com/watch?v=MxiWp8vkRFI After viewing, students first task is narrative essay (or flip book, or even better, an illustrated comic book) using the information from the two videos to tell the story of malaria’s life cycle, beginning with human infection from a mosquito bite, ending with transmission to a new human host. Then connect this to parts 1 and 2 of this lesson. What kind of ecological relationship does malaria have to humans (answer: parasitism)? How about its relation to mosquitoes (answer: commensalism)? Why? What kind of disease is malaria (answer: vector borne, tropical, protist-caused). Enrichment activity: using microscopes and water from our classroom aquaponic system, students examine protozoa and observe their behavior. Usually in our system there is an abundance of paramecia and amoebas, which can be seen feeding on bacteria and on algae. Form a vivid connection to malaria by asking students to imagine protozoa swimming in their blood, not pond water, and feeding on their red blood cells, not algae and bacteria. Standards: S8.B.1.1.3 Apply knowledge of characteristic structures to identify or categorize organisms (i.e., plants, animals, fungi, bacteria, and protista). S8.A.2.2.3 Describe ways technology (e.g., microscope, telescope, micrometer, hydraulics, barometer) extends and enhances human abilities for specific purposes. Objective: Sequence: Part 1: Review the basics of selection using this simple video from Bozeman: https://www.youtube.com/watch?v=3E4XZB6lKOE Provide a graph showing the frequency of peppered moth and how that frequency changed during industrialization. (Graph is in the Bozeman video and widely available on line). Provide a big blank for students to explain this frequency change. Ask students to use this word bank: variation, selection, predator, more frequent, less frequent, evolution. Now, explain that there are different types of natural selection (directional, disruptive, stabilizing). Watch this video together: Bozeman science video on types of selection: https://www.youtube.com/watch?v=W6eZ7zIIOas Watch this video up to minute 7. After that comes a discussion of sexual selection that can be left out here. Use a three columned organizer while watching: what are the three different kinds of natural selection? Explain and provide an example of each type. Part 2: First, watch this two minute video on how agriculture changed the ecology of malaria, creating selection pressure towards greater virulence: https://www.youtube.com/watch?v=64pvlCtH-Oo On the organizer, have students answer this question: why was malaria less virulent in the ancient past than it is now? How does this change connect to agriculture? Connect this to part one of the lesson: can we explain the emergence of the more virulent strain as a result of directional selection? Part 3: watch this simple video on the trade-off between virulence and transmissibility, with an emphasis on malaria: https://www.youtube.com/watch?v=GOghsFzYY8k Last question on organizer: why has malaria stopped growing more virulent? Have students explain its current level of virulence as a balance between costs and benefits. How can its current level of virulence be explained as a result of balancing selection, where less and more virulent strains are less fit (meaning, they have lower reproductive success)? Key vocabulary: evolution, virulence, transmissibility, trade-off hypothesis, directional selection, stabilizing selection, mutation, frequency, variation, cost-benefit analysis Standards: S8.B.2.1.1 Explain how inherited structures or behaviors help organisms survive and reproduce in different environments. S8.B.2.1.2 Explain how different adaptations in individuals of the same species may affect survivability or reproduction success. S8.B.2.1.3 Explain that mutations can alter a gene and are the original source of new variations. S8.B.2.2.2 Recognize that the gene is the basic unit of inheritance, that there are dominant and recessive genes, and that traits are inherited. S8.B.3.2.1 Use evidence to explain factors that affect changes in populations (e.g., deforestation, disease, land use, natural disaster, invasive species). S8.B.3.2.3 Describe the response of organisms to environmental changes (e.g., changes in climate, hibernation, migration, coloration) and how those changes affect survival. Objective: Students will learn that human beings have evolved in response to malaria. The mystery posed at the end of lesson 1 (why are almost all U.S. sickle cell disease victims African American?) will now be answered: sickle cell disease is the result of an adaptation to malaria. Students will understand that people with SCD are homozygous for the sickle cell allele, while people with SCT (sickle cell trait) are heterozygotes. Second, they will learn that evolution happens because of selection pressures, and that it also happens because of genetic drift. They will learn to recognize when evolution has occurred by studying the Hardy-Weinberg principle. They will use colored beans and a cup (the ‘mating chamber’) to model frequency change in alleles as a result of selection pressures (e.g., malaria), and as a result of drift. Note: this lesson adapts excellent materials developed by the Howard Hughes Medical Institute, including this worksheet: https://www.biointeractive.org/sites/default/files/Malaria_PopulationGenetics_Student.pdf My sequence is modeled on the one used in this HHMI worksheet, adapted to my 8th graders’ ability levels. Part 1: If necessary, review the basics of Mendelian genetics using Punnett squares. If W stands for the normal hemoglobin allele, and B for the sickle cell allele, have kids draw squares for the following matings: BB with BB, BW with BW, BW with BB, BW with WW. Pose this question: if the sickle cell allele is so harmful, why do so many Americans still have it? Answer: the sickle cell allele ‘hides’ in heterozygotes, meaning, heterozygotes are asymptomatic, so they can carry this allele with no fitness penalty, which in turn means that allele will persist in the population. Part 2: When populations are not evolving: Hardy-Weinberg equilibrium Watch this video from the Amoeba sisters: https://www.youtube.com/watch?v=7S4WMwesMts This video has a few simple problems which require students to use the H-W equation. Next, use the black and white beans to simulate evolution due to genetic drift. Assume the parent population has a genotype ration of 1:2:1, BB, BW and WW respectively, so you will have 5 parents who are BB, 10 who are BW and 5 who are WW. That means 30 beans altogether, 15 white and 15 black. Put 15 white beans and 15 black beans in the mating chamber. Pull out two beans at a time, record genotype, return beans to mating chamber. Continue this until you have 15 ‘offspring’. Record genotypes. Use math to calculate the percentage change in each genotype, and in each allele. There will almost surely be some difference in genotype frequency. Ask students whether this means evolution has occurred. Ask whether (and how) the results would be different, if the population were much larger (get them to see that frequency change is less when the population is large and compare this to sampling error with coin tosses. Kids know that if you toss a coin 10 times, you probably will not get heads exactly 5 times, but if you toss it a million times, the frequency of heads will be about 50 percent. Connect this to what we are doing with the black and white beans. Part 3: Simulate a selection pressure Watch this 15 minute video from HHMI: https://www.youtube.com/watch?v=Zsbhvl2nVNE&t=213s This video explains the sickle cell allele as an adaptation, a trait that becomes more frequent in a population because it gives a selective advantage. That advantage is malaria resistance. The sickle cell allele, when present in a heterozygote (someone who has one normal hemoglobin allele and one sickle cell allele) correlates with much lower parasite counts in malaria victims. We will use the beans to simulate the fitness benefits and costs of having this allele. Again use 30 beans, half B half W. Mix them in the mating chamber. Pick out a pair of beans. If BW or WW, mark it in the genotype chart and return the beans to the chamber. If the ‘baby’ is BB, however, return both beans to the mating chamber and do not mark anything on the chart. This simulates the 100 percent lethality of being homozygous for the sickle cell allele. After you have 15 offspring, calculate frequency change. Kids should see that the frequency of the B allele is going down fast. Now add a second generation to the experiment. Using the marks on your genotype chart, adjust the quantity of B and W beads in the mating chamber. The total number of B and W beads should no longer be 15 and 15. Let the quantity of each type be a function of how many BW and WW ‘babies’ you recorded on your genotype chart from the preceding experiment. Now, mate them again, with the same rules: BB’s get returned to the chamber without being recorded, but BW’s and WW’s get recorded and then returned. After you have 15 offspring, stop and calculate allele frequencies. Kids should again see a steep decline in the frequency of allele B. Help them see that this directional selection in action. Part 4: Heterozygote advantage: Reset the chamber to its original ratios, 15 B and 15 W. Again create a genotype chart with columns for BW and WW. Start making ‘babies,’ but this time, follow these rules: if a baby is BB, return the beads to the chamber and make no marks on the chart. If the baby is BW, make a hashmark on the chart. If the baby is WW, make a ‘half’ a hashmark on the chart. In other words, there is one hashmark in the WW column for every TWO WW babies that are born. This simulates two factors: BB is still 100 percent lethal, but WW is 50 percent lethal (because of greater susceptibility to malaria). Record frequency change in alleles. Another generation: Using the allele counts from the first generation, reset the mating chamber (note: this means that we do not start with 15 W and 15 B, but rather with the quantity of B and W that are on our genotype chart from the mating immediately above. Again, make 15 babies, using the same rules as above. Record frequency change. Ask students to compare the change here to the change we had when heterozygotes did not have an advantage. Heterozygote advantage maintains the frequency of the B allele in the population, and this explains the persistence of sickle cell disease in African American populations, despite the lethality of the BB homozygote. Here, then, is the answer to the mystery that began this lesson sequence: why is SCD only present in people of African descent? The answer is that SCD comes from an allele that is an adaptation to a parasite that is endemic to sub-Saharan Africa, and that this allele travelled to the U.S. in the genomes of enslaved Africans, who are the ancestors of today’s African Americans. Enrichment activity: Using the classroom aquaponic system, simulate a selection pressure in flowers. Students can grow a variety of flowers and then simulate a selection pressure by culling flowers of a certain type or aiding the reproduction of other types. This will change the frequency of the targeted trait in the next generation of flowers. This experiment would of course require many weeks to complete. Key vocabulary: Hardy-Weinberg principle, codominance, homozygote, heterozygote, adaptation, evolution, selection pressure, fitness, allele Standards: S8.B.2.1.1 Explain how inherited structures or behaviors help organisms survive and reproduce in different environments. S8.B.2.1.2 Explain how different adaptations in individuals of the same species may affect survivability or reproduction success. S8.B.2.1.3 Explain that mutations can alter a gene and are the original source of new variations. S8.A.1.3.1 Use ratio to describe change (e.g., percents, parts per million, grams per cubic centimeter, mechanical advantage). S8.A.2.1.3 Design a controlled experiment by specifying how the independent variables will be manipulated, how the dependent variable will be measured, and which variables will be held constant. Objective: Learn that resistance to insecticides is evolving in mosquitoes, just as resistance to anti-malarial medicines is evolving in the plasmodium itself. Learn that our use of pesticides to control disease vectors can have unintended and negative consequences, using bioaccumulation of DDT as a case study. Learn why malaria has been eliminated in the U.S. and in Europe but not in Africa. Students should synthesize these stories into one conclusion: that there is no ‘magic bullet’ solution to this disease, no inoculation or vaccine that can free humanity from this burden. The only lasting solution to the problem of malaria is to alter the way people live and work. In other words, the only solution to malaria is ecological, not biomedical. Part 1: Resistance Watch this 3 minute video on the evolution of resistance in mosquitoes: https://www.youtube.com/watch?v=D1aU7HNh4jM Here is a simple video on antibiotic resistance: https://www.youtube.com/watch?v=znnp-Ivj2ek Have a worksheet with this question: what explains the growth in resistance to pesticides? Can you connect this story to the decrease in effectiveness in antibiotics? Part 2: DDT and bioaccumulation Use this video from the Amoeba sisters on bioaccumulation, DDT and the bald eagle: https://www.youtube.com/watch?v=TZk6vcmLcKw Answer these questions: what are some of the unintended effects of pesticide use? Given what you now know about the evolution of resistance, do the benefits outweigh the costs? Part 3: Environmental ethics 1st Video: Rachel Carson and her ‘conservative’ critics: https://www.youtube.com/watch?v=ykPH56Udc60 Next questions on the worksheet: You have heard some criticisms of Rachel Carson, and of the ban on DDT. Do you think these criticisms are fair? Why or why not? Does it make sense to blame Carson for the ban on DDT, and thus hold her responsible for human deaths, given what we know about resistance to DDT among mosquitoes? 2nd video: Sonia Shah on why malaria persists in Africa: https://www.youtube.com/watch?v=SF242mxtZN8&t=2s Worksheet question: What does Sonia Shah say about the reasons why malaria has not died out in Africa? Shah says we “impose our own priorities on the people of the malarious world.” What does she mean? Why does she say the widespread use of pesticides (and now ITN’s, insecticide treated bed nets) are examples of this? What is social development, and how does a nation’s level of development affect its susceptibility to malaria? If America wants to fight malaria in Africa effectively, what should it do? Key vocabulary: mutation, variation, selection, resistance, bioaccumulation (biomagnifation), persistent chemicals, primary, secondary and tertiary consumers, food web, energy pyramid Standards: S8.A.1.2.1 Describe the positive and negative, intended and unintended, effects of specific scientific results or technological developments (e.g., air/space travel, genetic engineering, nuclear fission/fusion, artificial intelligence, lasers, organ transplants). S8.A.1.2.2 Identify environmental issues and explain their potential long-term health effects (e.g., pollution, pest controls, vaccinations). S8.B.3.3.1 Explain how human activities may affect local, regional, and global environments. S8.B.3.3.4 Explain the long-term effects of using integrated pest management (e.g., herbicides, natural predators, biogenetics) on the environment. S8.B.2.1.1 Explain how inherited structures or behaviors help organisms survive and reproduce in different environments. S8.B.2.1.2 Explain how different adaptations in individuals of the same species may affect survivability or reproduction success. S8.B.2.1.3 Explain that mutations can alter a gene and are the original source of new variations.Lesson 1: Sickle Cell Disease
Lesson 2: What is malaria?
Lesson 3: Evolution of malaria
Lesson 4: Human evolution in response to malaria: an introduction to population genetics
Lesson 5: Disease ecology, resistance, and public health citizenship
Note: This essay cites many documents produced by the World Health Organization (WHO). These documents do not identify a chief author; they are the collaborative work of dozens of writers and researchers. The WHO’s World Malaria Report, published annually, serves as my main source of statistics about malaria. I also use WHO news releases and websites, which do not identify a chief author. Allison, Anthony C. “The discovery of resistance to malaria of sickle‐cell heterozygotes.” Biochemistry and Molecular Biology Education, Nov. 2006. Barnwell, John. “Hepatic Kupffer Cells: The Portal That Permits Infection of Hepatocytes by Malarial Sporozoites?” Hepatology, May 2001. Carter, Richard. “Evolutionary and Historical Aspects of the Burden of Malaria.” Clinical Microbiology Reviews, Oct. 2002. Cyrklaff, Marek et al. “Hemoglobins S and C Interfere with Actin Remodeling in Plasmodium falciparum–Infected Erythrocytes.” Science, Dec. 2011. Fagan, Toby. “When was malaria first discovered and by whom?” Scientific American, July 17, 2000. Futuyma, Douglas. Evolutionary Biology, 3rd Edition, 1998: p. 385. Gardner, Malcolm et al. “Genome Sequence of the Human Malaria Parasite Plasmodium Falciparum.” Nature, Oct. 2002. Hume, Jennifer et al. “Human migration, mosquitoes and the evolution of Plasmodium falciparum.” Trends in Parasitology, March 2003. Loy, Dorothy et al. “Evolutionary history of human Plasmodium vivax revealed by genome-wide analyses of related ape parasites.” Proceedings of the National Academy of Sciences, Aug. 2018. Miller, Louis, et al. “The Resistance Factor to Plasmodium vivax in Blacks — The Duffy-Blood-Group Genotype, FyFy.” New England Journal of Medicine, Aug. 1976. Packard, Randall. Malaria: The Making of a Tropical Disease. Johns Hopkins Univ. Press, 2007. Sabeti, Pardas. “Natural Selection: Uncovering Mechanisms of Evolutionary Adaptation to Infectious Disease.” Nature Education, Jan. 2008. Webb, James. Humanity’s Burden: A Global History of Malaria. Cambridge University Press, 2009. Nobel Prize Biography of Alphonse Laveran: https://www.nobelprize.org/prizes/medicine/1907/laveran/biographical/ Podcast: “This podcast will kill you: Malaria”. http://thispodcastwillkillyou.com/2018/02/10/episode-8-abracadabra-go-away-malaria/ Kwiatowski, Dominic. “Malaria genomics: tracking a diverse and evolving parasite population.” Lecture on youtube: https://www.youtube.com/watch?v=bb1lgvx3HPg&t=295s National Center for Biotechnology Information website: https://www.ncbi.nlm.nih.gov/books/NBK2271/ National Institute of Health (NIH) website: https://ghr.nlm.nih.gov/condition/sickle-cell-disease#genesWorks Cited
Books and Articles
Videos, Podcasts and Websites